日本人はY染色体DNAのD系統を高頻度で持つ。
日本人の四割は縄文系です。
また、日本人の三割の弥生系も、O2b1aという日本固有の遺伝子です。
O2b1a = 彌生人の最大ハプルログル-ム.
トクシマで 28%, 大阪で 26% 発見されるなど関西で高く現われる.
(日本 23%, 韓国 8%, 中国 0%)
–
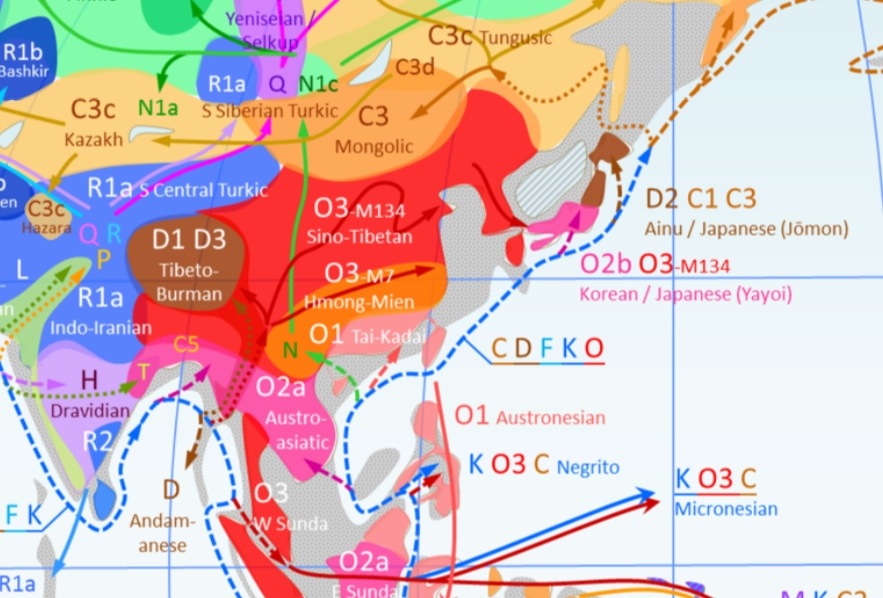
ハプログループDは、東アジアに住むためにアラビアから南アジアの沿海岸を通って東南アジアへ、さらに東南アジアから北方への経路を進んで移住していったと想定されるが、現在のインド・中国(漢民族)やその他の地域では全くその痕跡が見当らない。地球上で、ハプログループDが人口比に対して高頻度で見つかるのは日本、チベット、ヤオ族、アンダマン諸島、フィリピンマクタン島、グアム島、だけで、世界でも孤立した限られた地域でしか見つかっていない。しかし、これらのハプログループは、同じハプログループDに属していても、サブグループが異なるため、分岐してから数万年を経ていることを示している。またハプログループDは、中国、朝鮮、東南アジアにおいて一般的なハプログループOとはさらに遠く隔たっている。
ハプログループDは、今より約5万年前、アフリカにおいてハプログループDEから分岐し、沿海ルートを通ってアジアへ向かったと考えられている。今日、このハプログループDは、アジアの極めて限られた地域でしか見つかっていない。一方、同じくハプログループDEからわかれたハプログループEは、アフリカや中東、ヨーロッパなどで見つかっている。
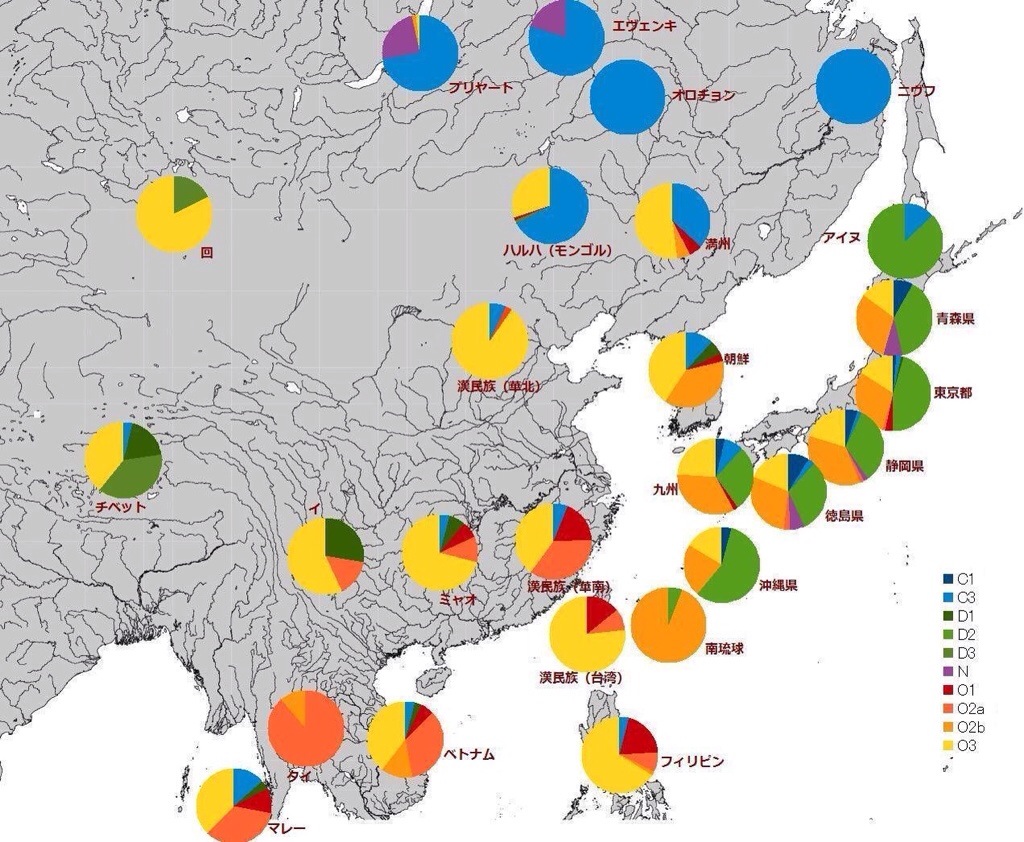
この系統は本土日本人・アイヌ・南北沖縄に固有に見られるタイプで、朝鮮半島や中国人にはほとんど見られないことも判明した。これは縄文人の血を色濃く残すとされるアイヌに88%見られることから、ハプログループD2は縄文人特有のY染色体だとされる。
アリゾナ大学のマイケル・F・ハマー (Michael F. Hammer) のY染色体分析でもYAPハプロタイプ(D系統)が扱われ、さらにチベット人も南北琉球同様50%の頻度でこのYAPハプロタイプを持っていることを根拠に、縄文人の祖先は約5万年前に中央アジアにいた集団が東進を続けた結果、約3万年前に北方ルートで北海道に到着したとする説を提出した。
現在世界でD系統は極めて稀な系統になっており、日本人が最大集積地点としてその希少な血を高頻度で受け継いでいる。それを最大とし、その他では遠く西に離れたチベット人、一部の台湾原住民に存続するだけである。これは、後に両者を隔てる広大な地域にアジア系O系統が広く流入し、島国日本や山岳チベットにのみD系統が残ったためと考えられている。
なお東西に引き離されたD系統は、長い年月により東(日本)がD2、西(チベット等)がD1、D3となった。D2系統は本土日本人・アイヌ・南北琉球の日本人集団固有であり他地域には希である。また、近年の遺伝子調査により、アイヌは縄文人の単純な子孫ではなく、オホーツク人等の北方民族と混血しており、複雑な過程を経て誕生したことが明らかになった[39]。ハプログループD2は本土日本人の平均でも、最も頻度が多く、またC1も他の国には見られないY染色体ハプログループとされる。
O2系統からは、他に東南アジアやインドの一部に見られるO2a系統と、日本のO2b1、中国東北部・朝鮮に多いO2bにそれぞれ分類される。そもそものO2b系統は長江周辺が発祥、ベトナム人に多い系統である。
崎谷満によれば、最初に日本列島に到達し、後期旧石器時代を担ったのはシベリアの狩猟民であるC3系統である(2 – 3万年前)。バイカル湖周辺からアムール川流域およびサハリンを経由して、最終氷期の海面低下により地続きとなっていた北海道に達した。また、一部はさらに南下し、北部九州に達した。崎谷は細石刃石器を用い、ナウマンゾウを狩っていたと考えている。
その後、約1万数千年前に、大陸からD2系統が入ってきた。これが縄文人である。D2は日本に多く見られる系統であり、アイヌ88%、沖縄県(北琉球の一部と南琉球)56%、本州42 – 56%で、東アジアでは存在しない。近縁のD1、D3がチベットで見られる。D系統は華北で東西に分かれ、東がD2、西がD1、D3になったと考えられる。
D2系統はアイヌ人88%、沖縄人56%、本土日本42~56%(参考までチベット33%)で、韓国ほぼ0%である。
D2の分岐は日本列島内で、氷河期の終わりと共に孤立したハプログループDの系統から独自にハプログループD2に分岐したと考えられる。
同じ頃、経路は不明であるが、インドに起源を持つC1系統が南九州に入ってきた。貝文土器を用い、縄文人とは異なる文化を南九州に築いた。
O1系統は台湾が起源である。崎谷満はオーストロネシア語族との関連があると想定している。台湾と近いにもかかわらず、日本列島ではO1はごく少数に過ぎない。
O2a/O2b系統について、崎谷満は長江文明の担い手だと考えている。O2b系統が移動を開始したのは約2800年前で、長江文明の衰退に伴い、O2aおよび一部のO2bは南下し、百越と呼ばれ、残りのO2bは西方及び北方へと渡り、日本列島、山東省、日本から朝鮮半島渡ったと崎谷満は主張している。長江文明の稲作を持ち込んだと考えられる
ハプログループD2は約3万年前に日本列島にて発生した型で日本以外ではみられないものである。
アイヌには南方系O系統は無く、完全なD系統であり、本土日本人と沖縄人には南方系のOが混ざっている。それでも本土日本人のD系統は半数以上である。
つまり事実上アイヌと本土日本人は大差がなく、アイヌに南方系を足したのが本土日本人となる。
結論として日本人は基本的に共通して、アイヌ人~本土日本人~沖縄人までD系統(北方系の古いモンゴロイド)がベースになっており、
日本人はY染色体DNAのD系統を高頻度で持つ事で有名である。世界的にもチベット人や中近東の人だけがこのDNAをもっている
東アジアの地域全体には南方系O系統が広く分布し、島国の日本や山岳のチベットにのみ古くからのD系統が残ったと考えられる
日本人は基本的に共通して、アイヌ人、本土日本人、沖縄人まで北方古モンゴロイドを基調としており、中国や韓国など東アジアの諸国と異なっている。
—————————————-
参考まで
—————————————-
遺伝子上(Y染色体ハプロタイプ)の状況
【 本土日本人 (%は混血比率)】
・DE系統D亜型D1B1/D1B2(約40~45% YAPあり)
・NO系統O亜型O2B1(約20~30%)
・NO系統O亜型O3(約10~16%)
※ 縄文人:D1B1/D1B2
※ 弥生人:O2B1(O3も含む可能性あり)
その他として数%「C1(日本固有)」「N」「C3」「O2B*」検出。
—————————————-
【 中国人(漢民族 %は混血比率)】←★DE系統なし
・CF系統C亜型C3 (約 8%)
・NO系統O亜型O1A(約10%)
・NO系統O亜型O2*(約10%)
・NO系統O亜型O2A(約 5%)
・NO系統O亜型O3 (約54%)←★中国人(漢民族)の主系統
※地方差あり
—————————————-
【 朝鮮人(韓民族 %は混血比率)】←★DE系統なし
・NO系統O亜型O2B*(約30%)
・NO系統O亜型O3(約45%)←★中国人(漢民族)の系統
・CF系統C亜型C3(約12%)
その他として、数%「D1A」「O1A」「N」「K」「D1B」「O2B1」が検出。
—————————————